
Los caballos y las emociones del rostro humano
Sep 24, 2017Este estudio presenta la primera evidencia de las habilidades de los caballos para discriminar espontáneamente entre expresiones faciales humanas positivas (felices) y negativas (enojadas) en fotografías. Estos resultados mostraron que las caras enojadas indujeron respuestas indicativas de una comprensión funcional de los estímulos: los caballos mostraron un sesgo de izquierda-mirada (una lateralización generalmente asociada con estímulos percibidos como negativos) y un aumento más rápido de la frecuencia cardíaca (FC) hacia estas fotografías.
Tales respuestas lateralizadas hacia la emoción humana han sido previamente documentadas solamente en perros, y los efectos de las expresiones faciales en la HR no se han demostrado en ningún estudio heterospecífico. Junto con las ideas que estos hallazgos proporcionan en la comunicación interespecífica, plantean interesantes preguntas acerca de la generalidad y adaptabilidad de la expresión emocional y la percepción entre especies.
1. Antecedentes
En muchas especies sociales, las emociones proporcionan valiosa información social y ambiental y es probable que desempeñen un papel clave para facilitar la cohesión y el funcionamiento del grupo.
Dado que la observación de la emoción positiva provoca el comportamiento de acercamiento hacia estímulos gratificantes, mientras que las emociones negativas promueven la evitación de posibles amenazas, la capacidad de respuesta a la emoción en otros es potencialmente altamente adaptativa.
La percepción de la emoción entre especies puede ser un reto donde la variación morfológica considerable divide el emisor y el receptor. Hasta la fecha, se ha encontrado que varias especies reconocen las expresiones emocionales humanas cuando se les presenta el conjunto completo de señales corporales, o después del entrenamiento para que se adapten específicamente a rasgos faciales asociados con emociones particulares.
Sin embargo, la medida en que las expresiones faciales pueden ser discernidas espontáneamente a través de las barreras de las especies ha recibido sorprendentemente poca atención.
Se ha realizado pruebas funcionalmente relevantes para explorar esto directamente, en un paradigma que nos permite evaluar tanto los mecanismos cognitivos y fisiológicos subyacentes involucrados.
La aparición de respuestas lateralizadas puede proporcionar un medio importante para evaluar cómo las señales se procesan cognitivamente. La valencia emocional percibida de los estímulos puede determinarse a través de sesgos hemisféricos, generalmente con especialización hemisférica derecha para procesar estímulos negativamente validados incluyendo encuentros agonísticos.
En consecuencia, también se ha informado de la especialización hemisférica izquierda para estímulos ambientales positivos, pero hay evidencia contradictoria sobre la lateralización de respuestas a situaciones sociales positivas.
La percepción lateralizada de las señales faciales heterospecíficas a la emoción sólo ha sido documentada en perros, donde las expresiones faciales humanas «enojadas» se ven con un sesgo de izquierda-mirada, aunque no se ve sesgos de mirada hacia expresiones faciales felices. Además, las medidas de la frecuencia cardiaca (FC) tienen el potencial de proporcionar ideas objetivas sobre la percepción de un animal de los estímulos externos. En los caballos, HR se correlaciona con los índices de comportamiento del estrés y fluctúa de acuerdo con el estrés del entrenador, lo que demuestra una potencial sensibilidad fisiológica a los afectados humanos.
Los caballos son un modelo ideal para la investigación de la comunicación interespecífica de la emoción, ya que son capaces de producir expresiones faciales complejas y percibirlas en especifico , así como ser sensible a las señales humanas, incluyendo señales faciales.
Además, las medidas de lateralización son particularmente apropiadas para los caballos dado sus ojos colocados lateralmente y su tendencia hacia los comportamientos lateralizados. Este estudio examina si los caballos discriminan espontáneamente y responden apropiadamente a expresiones faciales humanas positivas (felices) y negativas (enojadas). A cada caballo se le mostraron dos fotografías -una feliz y otra enojada- a lo largo de dos ensayos, mientras que se midieron sus respuestas conductuales y fisiológicas (HR).
Se esperaba que los estímulos negativos inducirían un comportamiento de evitación y un sesgo de la mirada de la izquierda, mientras que los estímulos positivos inducirían un comportamiento de aproximación y un sesgo de la mirada derecha o ningún sesgo. Además, se esperaba que los HRs de los caballos fueran más altos, para aumentar más rápido y requerir períodos de recuperación más largos en respuesta a estímulos negativos.
2. Material y métodos
Los caballos fueron reclutados en cinco caballerizas de Sussex y Surrey, Reino Unido, entre abril de 2014 y febrero de 2015.
La muestra final contenía 28 caballos en los análisis de comportamiento (21 caballos, siete yeguas, de 4 a 23 años, M = 15,46, sd = 5.25), con un subconjunto de 17 en los análisis de HR principales y 15 en los análisis de recuperación de HR.
Los estímulos eran fotografías de color laminadas de alta calidad de A3 (42 × 29,7 cm) de dos modelos montados en una cartulina A1, cada uno con una imagen positiva (feliz) y otra negativa ( figura 1a ). Las expresiones faciales fueron validadas utilizando descripciones del Sistema de Codificación de Acción Facial (FACS).
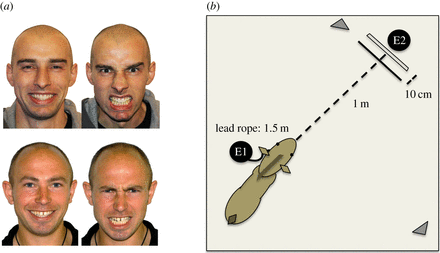
a ) Estímulos (L-R, positivos, negativos); (E1 = experimentador 1 que sostiene el caballo mientras se aleja del estímulo, E2 = experimentador 2 detrás del tablero, sosteniendo el estímulo, los triángulos representan las cámaras). Vea el material suplementario electrónico para la fotografía del montaje. (Versión en color en color.)
Los ensayos fueron conducidos en establos por un equipo de experimentadores.
En primer lugar, el experimentador 1 mantuvo el caballo sobre una cuerda de plomo suelta durante 4 minutos para obtener medidas de FC de línea de base, luego movió al caballo en su posición y esperó hasta que la FC volviera a la línea de base. El experimento 2 entró entonces en el establo y presentó los estímulos fotográficos en el campo de visión binocular del caballo.
La parte superior de la tabla se mantuvo a la altura de la marchita para estandarizar la colocación de los estímulos. Los estímulos se mantuvieron a 1 m de la nariz del caballo durante 10 s, luego se movieron hacia delante en 10 cm y se mantuvieron durante 10 s, luego volvieron a la posición original y se mantuvieron durante 10 s finales ( figura 1 b ).
Las mediciones de HR abarcaron el período de prueba más aproximadamente 5 s antes y después de la presentación del estímulo.
Durante las presentaciones, el experimentador 1 se colocó en el hombro izquierdo del caballo, lejos del estímulo, con el caballo sostenido en una cuerda de plomo suelta de 1,5 m (permitiendo el libre movimiento dentro de esta longitud) mientras que el experimentador 2 se agazapaba detrás del tablero de estímulos. Se controló la FC durante 4 minutos finales para evaluar las tasas de recuperación.
Cada caballo vio expresiones tanto positivas como negativas del modelo 1 o del modelo 2 con al menos dos meses de diferencia, contrapesadas igualmente por la emoción y el modelo. La HR se midió con un monitor Polar Equine ® RS800CX y se registraron ensayos con las videocámaras digitales Panasonic HC-X900 y HC-V720.
Las respuestas de comportamiento (duraciones de búsqueda, duración de aproximación y evitación y ocurrencias de comportamientos relacionados con el estrés: consulte el material complementario electrónico, tabla SI) fueron codificadas a ciegas a partir de videos en formato .mov en un Macbook Pro usando Sportscode Gamebreaker Plus ® 7.5.5 ( www.sportstec.com ).
Los experimentadores fueron ciegos al estímulo en el 82% de los ensayos y los análisis no mostraron diferencias en las respuestas de los sujetos de acuerdo a este factor. Diez de los 56 vídeos fueron codificados por AS y KG y se encontró que eran confiables en o por encima de r = 0,85, p ≤ 0,002.
3. Resultados
(a) Respuestas conductuales
El número de caballos que giraron inicialmente a la izquierda oa la derecha para cada emoción se evaluó mediante pruebas binomiales de dos colas.
Para cada sujeto, se calculó un índice de lateralidad (LI) para el tiempo total de observación: LI = ( L – R ) / ( L + M + R ), donde L , M y Rrepresentan la longitud de tiempo (s) , media y derecha, respectivamente. Las puntuaciones positivas indican un sesgo de la izquierda-mirada y las puntuaciones negativas un sesgo de la mirada derecha. Las desviaciones de la mirada binocular (nivel de probabilidad: 0) se midieron mediante pruebas t de una muestra (de dos colas).
Al ver estímulos negativos, más caballos parecían a la izquierda para su primer aspecto monocular que a la derecha ( n = 28, K = 20, p = 0.036). También hubo un sesgo de la izquierda-mirada en el tiempo de búsqueda total (índice de lateralidad) ( M = 0,23, sem = 0,07), t 27 = 3,49, p = 0,002. No hubo efectos de lateralidad en las respuestas a estímulos positivos ni en la primera mirada monocular ( n = 26, K = 16, p = 0,33), ni en el tiempo total de observación ( M = 0,09, sem = 0,063), t 27 = 1,48, p = 0,15 ( figura 2 ).
Figura 2.
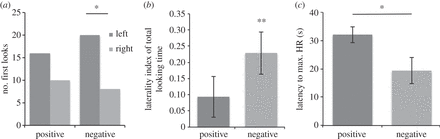
( a ) Frecuencias de la primera mirada por emoción; ( b ) índice de lateralidad media por emoción (± 1 sem); ( c ) tiempo mediano empleado para alcanzar la FC máxima durante la prueba (IC ± 95%). No hubo diferencias significativas en la duración de las observaciones cuando se compararon directamente las valencias, t 27 = -1,49, p = 0,15 (prueba t de muestras pareadas, dos colas). p > 0,05, ** p > 0,01.
No hubo diferencias significativas en la duración del enfoque frente a estímulos negativos (positivo, Mdn = 0,48, IC ± 1,58, negativo, Mdn = 0,00, CI ± 1,27), z = -0,97, p = 0,33, ni en la duración de la evitación , Mdn = 2,9, CI ± 1,23, positivo, Mdn = 1,14, CI ± 1,38), z = -1,03, p = 0,30 (pruebas de Wilcoxon, de dos colas).
Sin embargo, el tiempo que pasó mirando a la izquierda se correlacionó positivamente con el tiempo que se pasó evitando, r = 0,37, p = 0,005, pero no el tiempo que pasó acercándose, r = -0,12, p = 0,37, -0,003, p = 0,99, ni aproximación, r = 0,16, p = 0,23 ( ρ de Spearman).
Se observaron más comportamientos relacionados con el estrés en comparación con los estímulos positivos; sin embargo, el número de incidencias fue relativamente pequeño y los análisis estadísticos carecían de energía.
(b) Frecuencia cardiaca
Desde el comienzo de la fase de prueba, los HRs de los caballos aumentaron significativamente más rápido cuando se expusieron a estímulos negativos (Mdn = 19,4, CI ± 6,50) en comparación con los estímulos positivos (Mdn = 32,1, CI ± 4,01), z = -2,20, p = 0,028 ( figura 2) , prueba de Wilcoxon, de dos colas).
Sin embargo, el cambio promedio de HR de los caballos entre la línea de base y la prueba, la HR máxima absoluta y el tiempo de recuperación no fueron significativamente afectados por la emoción (cambio de HR: z = -0.54, p = 0.59; HR máxima: z = -0.26; p = 0.80; tiempo de recuperación: z = -1,22, p = 0,22, pruebas de Wilcoxon de dos colas, véase la tabla electrónica de materiales suplementarios SIV). Por último, la diferencia de HR media entre la prueba y la línea de base se correlacionó positivamente con el índice de lateralidad, r = 0,34, p = 0,047 y tiempo de evitación, r = 0,58, p > 0,001 ( ρ de Spearman).
4. Discusión
Los resultados conductuales y fisiológicos aquí presentados apoyan la hipótesis de que los caballos son capaces de reconocer y responder de una manera funcionalmente relevante a las expresiones faciales heterospecíficas (humanas) de ira.
Los caballos demostraron polarización hemisférica derecha hacia estímulos enojados (preferentemente viendo imágenes con el ojo izquierdo), que se correlacionaron positivamente con la duración de la evitación y el aumento medio en la HR; Además, los caballos mostraron un aumento general más rápido de la FC a la ira en comparación con los estímulos felices.
También hubo tendencias no significativas para realizar más conductas relacionadas con el estrés hacia estímulos enojados. Estos hallazgos plantean preguntas interesantes sobre la naturaleza del reconocimiento de la expresión emocional, incluyendo los roles relativos del aprendizaje y las habilidades innatas en su desarrollo.
Existen numerosas explicaciones posibles para la aparición de las habilidades de los caballos para discriminar las expresiones faciales humanas particulares.
Los caballos pueden haber adaptado una capacidad preexistente (ancestral) para responder apropiadamente a las expresiones emocionales negativas de los congéneres y, a través de su coevolución con los humanos, transfirieron esta habilidad a una especie morfológicamente diferente.
Alternativamente, los individuos pueden tener que aprender a interpretar las expresiones humanas durante su experiencia de vida con los seres humanos.
En apoyo de este último argumento, la familiaridad se encuentra ser un factor significativo en el reconocimiento de los perros de las expresiones humanas; se desempeñan mejor cuando se enfrentan con sus dueños o con personas del mismo sexo que sus dueños, lo que sugiere que la experiencia de vida tiene un papel importante en la formación de esta capacidad. Para elucidar los mecanismos evolutivos y ontogenéticos involucrados, las respuestas de las especies y los individuos con diferentes grados de exposición humana deben ser comparados.
El sesgo hemisférico derecho de los caballos hacia las expresiones faciales negativas del hombre se esperaba sobre la base de la literatura anterior y también se observa en perros.
Sin embargo, la falta de una respuesta lateralizada a las expresiones positivas en nuestro estudio es más difícil de interpretar. Esto también se observa en los perros y puede ser porque los estímulos positivos son menos evidentes.
El reconocimiento de los estímulos negativos tiene una relevancia funcional particular, ya que permite a los individuos anticipar posibles consecuencias negativas (por ejemplo, manipulación brusca, castigo).
Alternativamente, en este estudio, los caballos pueden no percibir los estímulos como abiertamente positivos debido a la falta de familiaridad de los seres humanos representados en los estímulos y de la puesta en marcha experimental.
También es notable que en nuestros experimentos, los estímulos fotográficos eran de dos machos desconocidos. La generalidad de nuestros hallazgos podría ser investigada más a fondo a través de estudios futuros sobre los efectos particulares de la identidad, la familiaridad, la edad y el género sobre las habilidades para discriminar una gama de expresiones emocionales.
5. Conclusión
Aquí presentamos la primera evidencia de la habilidad de los caballos de discriminar espontáneamente, tanto de manera conductual como fisiológica, entre expresiones faciales humanas positivas y negativas, y la primera evidencia de expresiones faciales heterospecíficas de emoción que afectan a la HR de una especie.
Esto plantea preguntas intrigantes acerca de la flexibilidad y adaptabilidad de la percepción emocional en este contexto, el rol potencial de la experiencia y la posibilidad de una capacidad generalizable, conservada y extendida para leer señales emocionales a través de las especies.
Ética
Esta investigación se adhiere a las directrices de la Asociación para el Estudio de Comportamiento Animal (ASAB) y fue aprobada por el Comité de Revisión Ética de la Universidad de Sussex (ERC), número de referencia: Non-ASPA 3-January 14. Se obtuvo el consentimiento por escrito de ambos modelos presentados en los estímulos ( figura 1a ).
No hubo diferencias significativas en la duración del enfoque frente a estímulos negativos (positivo, Mdn = 0,48, IC ± 1,58, negativo, Mdn = 0,00, CI ± 1,27), z = -0,97, p = 0,33, ni en la duración de la evitación , Mdn = 2,9, CI ± 1,23, positivo, Mdn = 1,14, CI ± 1,38), z = -1,03, p = 0,30 (pruebas de Wilcoxon, de dos colas). Sin embargo, el tiempo que pasó mirando a la izquierda se correlacionó positivamente con el tiempo que se pasó evitando, r = 0,37, p = 0,005, pero no el tiempo que pasó acercándose, r = -0,12, p = 0,37, -0,003, p = 0,99, ni aproximación, r = 0,16, p = 0,23 ( ρ de Spearman).

